Enhance your research with PE-Cy7 alternative with reduced spillover and monocyte background! Try BD Horizon RealBlue™ 780 SKU# [755602] today!
More Info on RB780


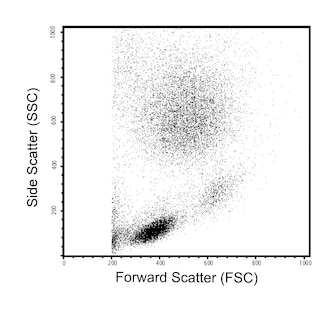
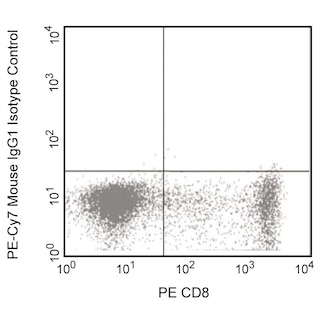

For Research Use Only. Not for use in diagnostic or therapeutic procedures.